Gissell Estrada-Rodríguez (DMAT, CRM)
 Mathematical modelling of biological, physical, and social phenomena using partial differential equations (PDEs) to describe the average behaviour of underlying microscopic (individual-level) dynamics has received significant attention [6, 11]. Since A. Turing’s foundational work on reaction–diffusion systems, followed by the influential chemotaxis model by Patlak, Keller, and Segel in the 1950s [13, 16], PDE models have become essential tools in understanding self-organisation.
Mathematical modelling of biological, physical, and social phenomena using partial differential equations (PDEs) to describe the average behaviour of underlying microscopic (individual-level) dynamics has received significant attention [6, 11]. Since A. Turing’s foundational work on reaction–diffusion systems, followed by the influential chemotaxis model by Patlak, Keller, and Segel in the 1950s [13, 16], PDE models have become essential tools in understanding self-organisation.
Despite their success, PDEs are often constructed directly at the continuum level; interactions are typically based on phenomenological considerations at the population scale. This top-down approach can limit their ability to capture micromechanical behaviours and complicates validation against experimental data. A concrete illustration appears with the classical Keller–Segel model for chemotaxis: while rigorous derivations from stochastic particle systems exist [18], recent work [10] suggests an alternative microscopic mechanism («resident–explorer» interactions) producing analogous aggregation patterns. Establishing robust links between PDE models and their microscopic counterparts enhances predictive power and makes macroscopic models more valuable for experimental data analysis.
In collective cell migration, these limitations arise naturally. Cell–cell adhesion, a fundamental mechanism during tissue formation, has been studied using coupled nonlinear nonlocal PDE systems for two populations of cells [3, 9, 14], where a mix of nonlocal adhesion and local saturation captures biologically relevant behaviours. Nonlinear diffusion (short-range repulsion) plus nonlocal aggregation imposes analytical and numerical challenges, especially under realistic conditions. Interaction parameters require careful calibration and are hard to measure experimentally. In addition, identifying biologically relevant interaction potentials remains a difficult task. Numerical results in [3] show such PDE systems qualitatively capturing experimental outcomes under different adhesion hypotheses, see Figure 3. Data-driven identification of interaction potentials [4] and efficient parameter estimation [2] can further refine these descriptions.
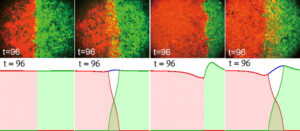
Figure 3: Top row: Cell-cell adhesion experiments from [12]. Bottom row: Numerical results from [3] using the system (7)
When mathematical models are improved using biological data, they accelerate discovery. Yet experimental validation of refined hypotheses is a bottleneck due to the time, cost, and complexity of biological experiments. To address this, we propose robotic swarms as controlled, measurable model systems for testing biological hypotheses. Compared to purely computational simulations, robot swarms introduce natural noise (e.g., sensing/actuation delays, limited communication), analogous to biological limitations, and may exhibit emergent behaviour (jamming, fragmentation) not predicted by simulations. This offers a realistic and flexible platform to speed the cycle of prediction, testing, and refinement, bridging modelling and experimental validation.
In this context, the present work aims to develop a comprehensive analytical, numerical, and experimental framework for studying nonlocal PDEs with biologically relevant features such as collective cell migration and adhesion. Central to this approach is the use of programmable robotic swarms as controllable physical testbeds for validating and refining PDE-based models. By integrating mean-field limit theory, data-driven parameter estimation, and asymptotic-preserving numerical schemes with systematically designed robot experiments, this framework enables direct testing of movement hypotheses under realistic conditions that include noise and emergent effects. Two principal objectives guide this effort: (i) to perform a qualitative comparison between PDE solutions and observed robotic swarm behaviours, with particular emphasis on parameter inference, and (ii) to establish rigorous error bounds that quantify discrepancies between microscopic and macroscopic descriptions, thereby characterising finite-size effects inherent to experimental systems.
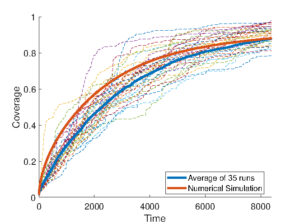
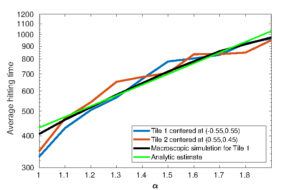
Figure 4: Top: Comparison of area coverage from a nonlocal PDE with averaged robotic simulations. Bottom: Comparison of hitting times. All plots correspond to 5 robots. Taken from [7]
Previous works [7, 8] derived nonlocal macroscopic PDEs for individual robot movement from detailed models of local communications and interactions, identifying limiting equations across temporal/spatial scales (see Figure 4). The combination of mathematical tools and robotic systems opens a zoo of questions between mathematical biology, analysis, and optimisation. For instance, spontaneous cell organisation leading to shape formation (morphogenesis) has been recently studied using a swarm of robots [5, 17]. In these works, robots exhibited collective behaviour by following classical reaction-diffusion PDEs à la Turing, relying entirely on local interactions with their neighbours. Differential adhesion hypothesis to program robot swarms was explored in [15], where no mathematical model was used, only simple rules mimicking cell behaviour. The use of tailored PDE models for cell adhesion provides a rigorous framework to scale swarm behaviours across system sizes and it enables efficient inference of key parameters such as adhesion strength and sensing radius directly from data.
PDE solutions vs. robotic swarms for cell–cell adhesion. Cell–cell adhesion involves binding through surface proteins. Although well studied theoretically, its quantitative calibration remains limited by scarce biological data. We therefore propose to calibrate the following two-population adhesion PDE system:
\begin{equation}
\begin{aligned}
\partial_t \rho_1&=\nabla\cdot\Bigl[\rho_1\nabla\Bigl(b_1(\rho_1+\rho_2)+ W_{11}*\rho_1+W_{12}*\rho_2\Bigr) +\Sigma_1\nabla\rho_1\Bigr]\ ,\\
\partial_t \rho_2&=\nabla\cdot\Bigl[\rho_2\nabla\Bigl(b_2(\rho_1+\rho_2)+ W_{22}*\rho_2+W_{21}*\rho_1\Bigr)+\Sigma_2\nabla\rho_2\Bigr]\ , \;\;\;\; (7)
\end{aligned}
\end{equation}
posed in a \(2\)-dimensional bounded domain with smooth boundary conditions. The calibration is performed using data collected from robotic experiments—designing two robot populations with different neighbour-to-neighbour communication strengths and sensing distances, which naturally map to adhesion and interaction radii. Here \(\rho_i(x,t)\), \(i=1,2\), are the population densities of each cell-type, \(b_i,\Sigma_i>0\) are the nonlinear and linear diffusion coefficients, respectively, and \(W_{ij}\) are the interaction potentials. The parameters we seek to estimate include the nonlinear diffusion coefficient and the coefficients governing the attractive and repulsive forces within \(W_{ij}\).
As a first step, we will perform a qualitative comparison between the PDE numerics and the observed robotic configurations, extending area-coverage metrics previously used for single-population settings [7]. Parameter refinement will follow a hybrid approach combining a finite number of trajectories with PDE descriptions [2]. The advantage of our approach in [2] lies in combining two key elements: a finite number of individual trajectories, for which we have accurate biological/robotic data, and the use of PDE descriptions, for which we have efficient parameter estimation schemes. This combination makes parameter estimation more efficient and computationally feasible compared to individual-based models, which require all trajectories for calibration. Beyond fixed-form (e.g., Morse) potentials, we will test multiple interaction kernels and nonlinear diffusions at both experimental and PDE levels.
Finite-size effects in PDE descriptions. While studying collective behaviour, a major difficulty arises when trying to quantitatively measure how well the robots swarm achieved a prescribed spatial configuration, since often the target density is given as a continuous function and the robots swarm is actually composed of a finite number of individuals at discrete locations. These finite size effects are inherent in continuum PDE models, which constitute a problem when comparing PDE descriptions with individual particle simulations, as corroborated in [20]. Robotic systems are often small, compared to their biological counterparts, and finite size effects are measurable.
The validity of the continuum limit as the number of particles tend to infinity for weak interactions, an unrealistic scenario in robotics and cell-cell interacting systems, was established in [18]. Preliminary work towards the derivation of rigorous error bounds on the discrepancy between microscopic and macroscopic models, which depend on the swarm population size and other physical characteristics such as sensing radius, was derived and validated in a series of papers by A. Bertozzi et al. [1, 19, 21] for a swarm of robots in the absence of interactions. Our aim is to extend the previous analytic framework to include (i) specular-type interactions between individuals of the same population, and ultimately, between individuals of different populations, (ii) different sensing radii depending on the different species. As a first step, a robot population will perform a simple task of area coverage where robot-to-robot interactions are considered. A second step will include two populations adhesion models.
Let us start by considering a bounded region \(\Omega\in \mathbb{R}^d\), the desired robots distribution \(\rho(z)\) in \(\Omega\) and \(N\) the number of robots with positions \(x_1,…,x_N\) in \(\Omega\). To describe the discrete positions of the robots A. Bertozzi et al. introduced a blob function \(B_\varepsilon(z)=\varepsilon^{-d}K(z/\varepsilon)\), which is normalised to \(1\) and \(\varepsilon>0\) describes the sensing radius. This is analogous to the purely repulsive potential introduced in [2] to describe nonlinear diffusion for the case of cell-cell interactions. The function \(K(\cdot)\) is generally assumed to be a Gaussian, namely, the robot is more efficient at its task locally than at larger distances from its current position. Then, if \(\varepsilon\) is small, we can derive a continuous distribution of the discrete positions of the robots given by \(\rho_\varepsilon^N(z)\simeq\frac{1}{N}\sum_{i=1}^N B_\varepsilon(z-x_i)\), which is analogous to the so-called empirical measure (\(\rho^N(x)=\frac{1}{N}\sum_{i=1}^N\delta_{x_i}(x)\)) when \(\varepsilon\to 0\). The error metric is naturally given now by $$e^\varepsilon_N(x_1,…,x_N)=\int_\Omega|\rho^N_\varepsilon(z)-\rho(z)|dz\ ,$$ which converges to zero as \(N\to\infty\).
Building on these ideas, a quantitative measure is introduced to characterise finite-size effects in the presence of strong interactions, which can provide a formal mathematical validation of the swarm control strategies, allowing us to determine the required number of robots and optimal robot sensing radius to achieve a final prescribed configuration. Gradient based optimisation algorithms will be used to minimize the error between the target distribution \(\rho(z)\), and the positions of the robots given by \(\rho_\varepsilon^N(z)\), given \(\rho\), \(N\) and \(\varepsilon\).
References
[1] B. G. Anderson, E. Loeser, M. Gee, F. Ren, S. Biswas, O. Turanova, M. Haberland, and A. L. Bertozzi. Quantitative assessment of robotic swarm coverage. arXiv:1806.02488, 2018.
[2] J. A. Carrillo, G. Estrada-Rodriguez, and S. Gomes. Parameter estimation for cell-cell adhesion dynamical models. In preparation, available upon request, 2025.
[3] J. A. Carrillo, H. Murakawa, M. Sato, H. Togashi, and O. Trush. A population dynamics model of cell-cell adhesion incorporating population pressure and density saturation. Journal of Theoretical Biology, 474:14–24, 2019.
[4] J. Carrillo de la Plata, G. Estrada-Rodriguez, L. Mikolás, and S. Tang. Sparse identification of nonlocal interaction kernels in nonlinear gradient flow equations via partial inversion. Mathematical Models and Methods in Applied Sciences, 35(05):1073–1131, 2025.
[5] D. Carrillo-Zapata, J. Sharpe, A. F. T. Winfield, L. Giuggioli, and S. Hauert. Toward controllable morphogenesis in large robot swarms. IEEE Robotics and Automation Letters, 4(4):3386–3393, 2019.
[6] L. Chen, K. Painter, C. Surulescu, and A. Zhigun. Mathematical models for cell migration: a non-local perspective. Philosophical Transactions of the Royal Society B, 375(1807):20190379, 2020.
[7] S. Duncan, G. Estrada-Rodriguez, J. Stocek, M. Dragone, P. A. Vargas, and H. Gimperlein. Efficient quantitative assessment of robot swarms: coverage and targeting lévy strategies. Bioinspiration & Biomimetics, 17(3):036006, 2022.
[8] G. Estrada-Rodriguez and H. Gimperlein. Interacting particles with levy strategies: limits of transport equations for swarm robotic systems. SIAM Journal on Applied Mathematics, 80(1):476–498, 2020.
[9] C. Falcó, R. E. Baker, and J. A. Carrillo. A local continuum model of cell-cell adhesion. SIAM Journal on Applied Mathematics, 84(3):S17–S42, 2023.
[10] T. Funaki, H. Izuhara, M. Mimura, and C. Urabe. A link between microscopic and macroscopic models of self-organized aggregation. Networks and Heterogeneous Media, 7(4):705–740, 2012.
[11] D. Horstmann. From 1970 until present: the keller-segel model in chemotaxis and its consequences. 2003.
[12] S. Katsunuma, H. Honda, T. Shinoda, Y. Ishimoto, T. Miyata, H. Kiyonari, T. Abe, K.-i. Nibu, Y. Takai, and H. Togashi. Synergistic action of nectins and cadherins generates the mosaic cellular pattern of the olfactory epithelium. Journal of Cell Biology, 212(5):561–575, 2016.
[13] E. F. Keller and L. A. Segel. Initiation of slime mold aggregation viewed as an instability. Journal of Theoretical Biology, 26(3):399–415, 1970.
[14] H. Murakawa and H. Togashi. Continuous models for cell–cell adhesion. Journal of Theoretical Biology, 374:1–12, 2015.
[15] M. Pan, Y. Yang, X. Qin, G. Li, N. Xi, M. Long, L. Jiang, T. Zhao, and L. Liu. Applying the intrinsic principle of cell collectives to program robot swarms. Cell Reports Physical Science, 5(8), 2024.
[16] C. S. Patlak. Random walk with persistence and external bias. The Bulletin of Mathematical Biophysics, 15(3):311–338, 1953.
[17] I. Slavkov, D. Carrillo-Zapata, N. Carranza, X. Diego, F. Jansson, J. Kaandorp, S. Hauert, and J. Sharpe. Morphogenesis in robot swarms. Science Robotics, 3(25):eaau9178, 2018.
[18] A. Stevens. The derivation of chemotaxis equations as limit dynamics of moderately interacting stochastic many-particle systems. SIAM Journal on Applied Mathematics, 61(1):183–212, 2000.
[19] C. Wang, Y. Zhang, A. L. Bertozzi, and M. B. Short. A stochastic-statistical residential burglary model with finite size effects. In Active Particles, Volume 2, pages 245–274. Springer, 2019.
[20] L. Wang and B. Yan. An asymptotic-preserving scheme for the kinetic equation with anisotropic scattering: Heavy tail equilibrium and degenerate collision frequency. SIAM Journal on Scientific Computing, 41(1):A422–A451, 2019.
[21] F. Zhang, A. L. Bertozzi, K. Elamvazhuthi, and S. Berman. Performance bounds on spatial coverage tasks by stochastic robotic swarms. IEEE Transactions on Automatic Control, 63(6):1563–1578, 2017.